教員と研究テーマ

魚類・両生類の多様な環境適応メカニズム~生理機構とその進化~
脊椎動物(特に、魚類や両生類)が、海、淡水、陸上という多様な環境に適応するための仕組みを、遺伝子から細胞、個体、進化の幅広い視点から解明することを目指しています。
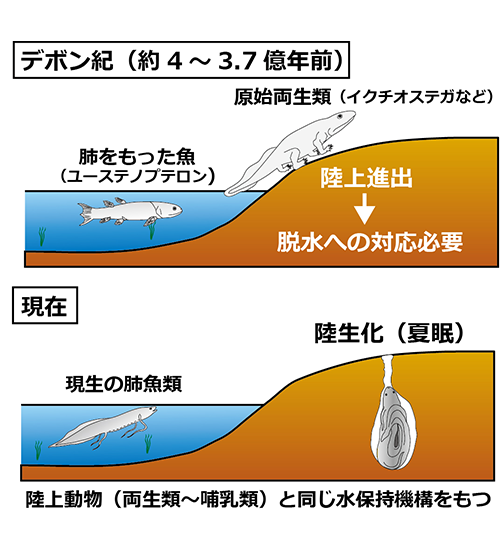
今野研究室では、生命維持の基本条件のひとつである ホメオスタシス(恒常性の維持)を研究の軸として、脊椎動物、特に、魚類や両生類がどのようにして多様な環境(海水、淡水、乾燥地域など)に適応しているのかを、生理メカニズムやホルモンの作用から探っています。また、四肢動物(両生類~哺乳類)の祖先は、その進化の過程で水から陸上へと進出してきましたが、両生類の祖先はいつ、どのような仕組みを獲得したことで上陸できたのでしょうか。進化的に両生類に最も近い魚である肺魚の生理機構を調べることで、その疑問に対する答えを探っています。
このような研究を通じて、環境の変化に対する動物の巧みな生き方や進化の不思議など、ホメオスタシスの統合的な理解を目指しています。

広塩性と狭塩性の違いを生むメカニズム
川(淡水)と海を往来するサケやウナギは、異なる浸透圧環境に、どのような仕組みを働かせて適応しているのでしょうか?淡水に生息するコイやキンギョ(狭塩性魚)は海水中で生きることができませんが、同じ淡水魚のメダカ(広塩性魚)は海水にも適応して繁殖することさえできます。同じ淡水魚でも、どうしてこのような違いがあるのでしょうか?この狭塩性と広塩性の違いには、鰓や腎臓といった体液浸透圧を調節する器官の構造的な違いだけでなく、さまざまなホルモンの働きやイオン輸送体、それらの発現を制御する転写因子が関係していると考えられていますが、未だその仕組みの全容は明らかになっていません。
私たちは、メダカの鰓に発現する浸透圧ストレス転写因子(Ostf1)やカルシウム活性化クロライドイオンチャネル(ANO1)に注目して、これらがメダカの海水適応や、狭塩性と広塩性の違いにどのように関わっているのかを調べています。

脊椎動物の水生から陸生への進化を支えた生理機構
私たちの遠い祖先は、かつて水中に住み、ヒレをもった魚から、手足を備えた両生類へと進化してきました。しかし、原始両生類が陸上に適応するためにはさまざまな生理機構を水生型から陸生型へと変化させる必要がありました。魚類でありながら、肺呼吸をして、陸上でも生存できる、“肺魚”という魚に注目して、魚類がどのようにして陸上へと適応できたのか?という進化の謎の解明に取り組んでいます。
● 夏眠の仕組みの解明
肺魚は、雨季には他の魚類と同様に水中で生活していますが、川の水が干上がる乾季になると地中に潜り、繭を作って、“夏眠”という乾燥に対する適応行動をとります。夏眠では、何も飲むことも食べることもできませんが、半年や数年間も生き延びることができます。長期の絶食状態であるにも関わらず生命を維持するためには一体どのような仕組みが働いているのでしょうか?私たちは、水中の肺魚と夏眠した肺魚を用いたさまざまな解析から、夏眠の仕組みや、長期の生命維持に働く生理機構について調べています。
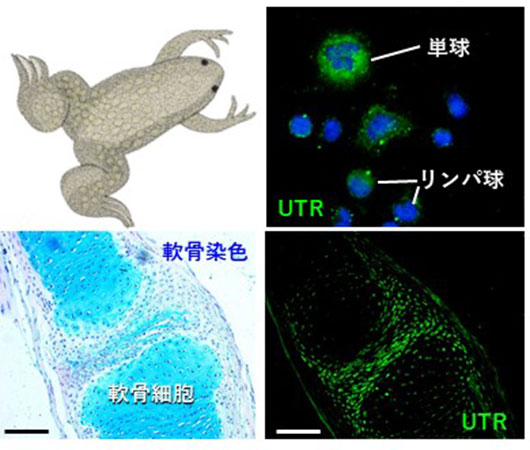
魚類、両生類から新しいホルモン機能を探す
魚類の尾部下垂体から発見されたウロテンシンⅡというホルモンは、強力な血管収縮作用を示すことが知られています。私たちは、ウロテンシンⅡ受容体がツメガエルの白血球(単球やリンパ球)や軟骨細胞に発現することを発見し、新たなホルモン作用の可能性を見出しました。その後の研究で、ウロテンシンⅡは白血球でのサイトカイン合成や軟骨基質形成作用をもつことが示唆されています。これらの作用は、これまでに報告がされていない新しいホルモン機能であると考えられ、応用研究も視野に入れて研究を行っています。
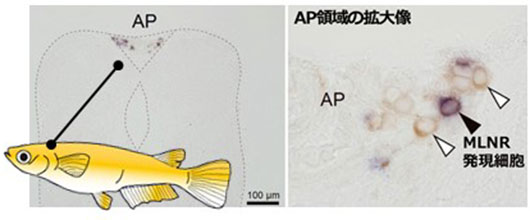
消化管(腸)から分泌されるモチリンというホルモンは、哺乳類では空腹時の胃腸管収縮に作用します。わかりやすくいうと、お腹が空いて「グ~」となるのは、モチリンが分泌されて腸のぜん動運動を促進させているからです。最近、メダカを用いた私たちの研究によって、モチリンが脳の延髄最後野や間腎腺(副腎)のカテコラミン産生細胞に作用していることが明らかとなりました。これは哺乳類でもまだ報告されていない発見であり、現在、その新しいホルモン機能について調べています。