研究トピックス TOPICS
- ホーム
- 研究トピックス一覧
- 生物学科の研究トピックス一覧
- 研究トピックス
新規のゴマリグナン合成酵素遺伝子の同定
【生物学科】山本 将之
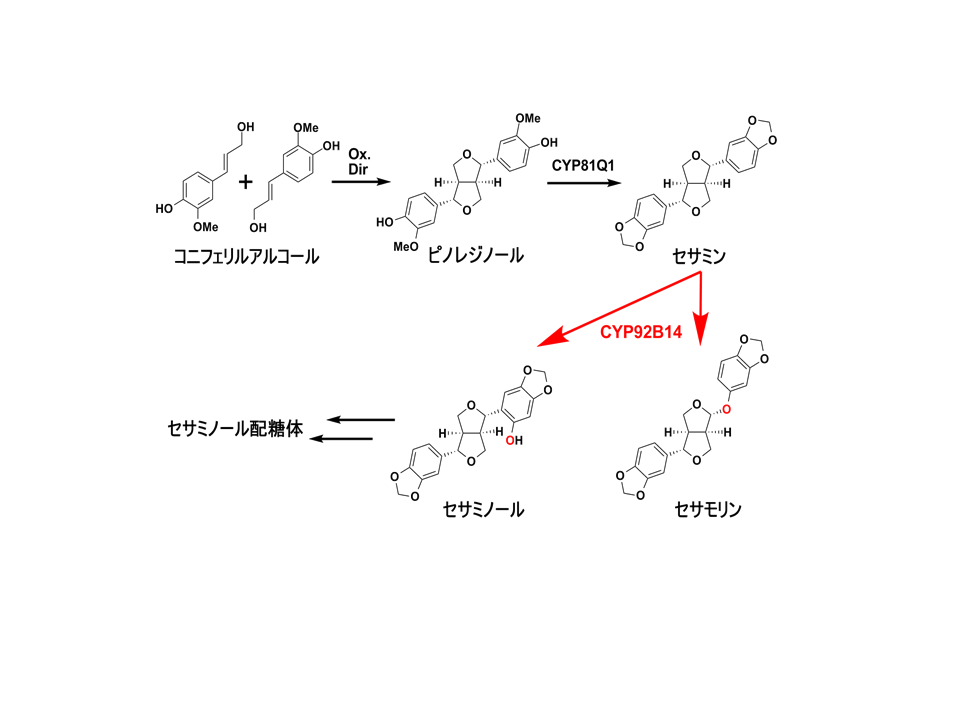
ゴマは種子中に多量の油脂を含む重要な油糧作物です。ゴマ種子中にはそのほかにも、リグナンと呼ばれる物質が含まれています。ゴマが産生するリグナン(ゴマリグナン)はいくつか種類がありますが、主にセサミン、セサモリン、セサミノールの3種類が種子中に豊富に存在します(図1)。最近、セサミンをはじめとしたゴマリグナンが持つ健康機能性に注目が集まり、多くの研究がなされています。しかし、ゴマリグナンがゴマの種子中でどのように合成されるかについては不明な点が多く、セサミンの生合成酵素 (CYP81Q1: Ono et al., 2006) が明らかになっているに過ぎませんでした。
富山大学理学部では様々なゴマ系統を維持しています。これらのゴマ系統についてのゴマリグナンの分析から、セサモリンをほとんど含まない系統(セサモリン低含有系統)が見出されています。この系統ではセサモリン生合成酵素が変異していると考え、セサモリン低含有の原因遺伝子の同定を行うことで、セサモリン合成酵素が同定できると考えました。
まずセサモリン低含有系統と、通常のセサモリン含有系統を交配しF6世代の遺伝集団を作成しました。F6集団160個体の種子中のセサモリン含有率(セサモリン含有量/セサミン、セサモリン、セサミノールの含有量の和)を調査したところ、5%以下の低含有個体と、15-45%の含有個体が1:1の割合で分かれ(図2)、セサモリン含有形質が1つの遺伝子座により決定されている可能性が高いことが分かりました。
続いて、F6集団の各個体からDNAを調製し、セサモリン含有形質と連鎖するDNAマーカーの探索を行いました。その結果、連鎖する2つのマーカー(90036と86884)が得られ、このうち90036はセサモリン含有形質と非常に強く連鎖していることが示されました(図3)。90036付近に存在する遺伝子を探したところ、セサミン合成酵素CYP81Q1と同じグループに属する遺伝子(シトクロムP450遺伝子)がいくつか見出されました。これらの発現の調査、塩基配列の比較解析を行ったところ、種子で発現している遺伝子 の1つ(CYP92B14) がセサモリン低含有系統では1塩基の挿入によって、指令するアミノ酸配列が短くなっていることが示されました(図4)。
通常のタイプ(セサモリン含有系統型)のCYP92B14を酵母で発現させ、セサモリン合成するかどうか調べたところ、この酵素はセサミンからセサモリンを生成することが明らかになりました。また、この酵素はセサミンからセサミノールを生成する活性も持つことが分かりました(図1)。
以上の結果から、セサモリン低含有形質の原因遺伝子CYP92B14がセサミンからセサモリンとセサミノールを生成する新規のゴマリグナン合成酵素遺伝子であることが分かりました。
この研究は、公益財団法人サントリー生命科学財団、サントリーグローバルイノベーションセンター(株)、龍谷大学、神戸大学と共同で行いました。
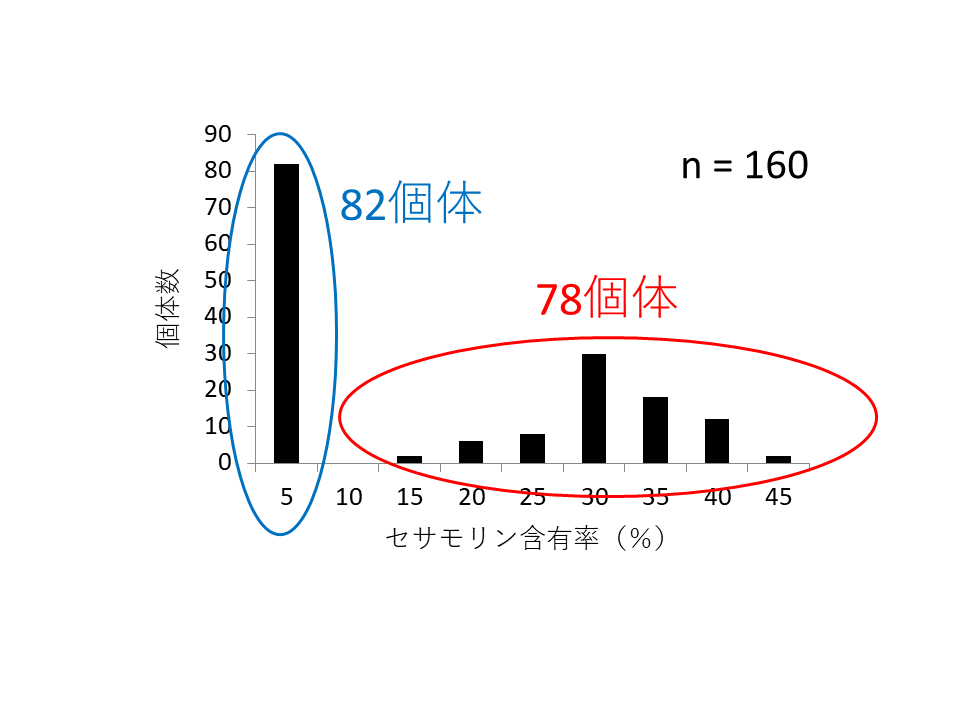
セサモリン低含有個体と、含有個体がおよそ1:1の割合で分かれた。
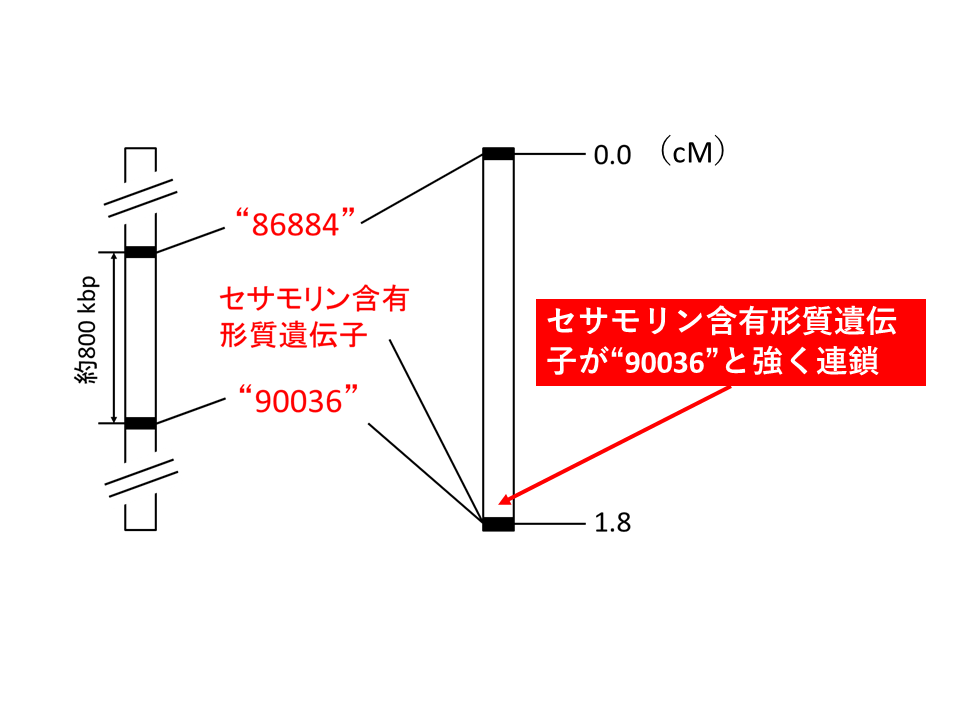
左は物理地図(DNAの長さに基づく地図)、右は連鎖地図(遺伝子座間の組換え頻度に基づく地図)を表している。
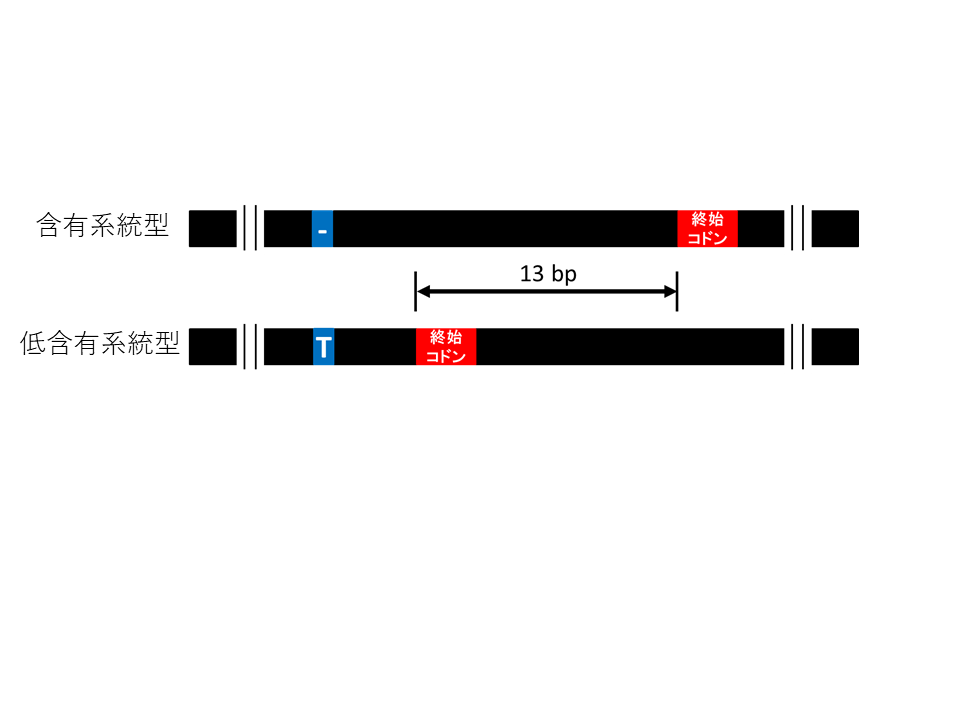
セサモリン含有個体と低含有個体のCYP92B14の塩基配列を示した。低含有個体型では含有個体型に比べて、1塩基の挿入がおこっており、この結果終止コドンの出現位置が早まることが分かった。
プレスリリース (2017/12/18:大学院理工学研究部(理学)の山本将之講師らの研究グループが新規のゴマリグナン生合成酵素遺伝子を世界で初めて同定)