研究トピックス TOPICS
- ホーム
- 研究トピックス一覧
- 生物学科の研究トピックス一覧
- 研究トピックス
日本のタンポポの倍数性
【生物学科】岩坪 美兼
道端、公園などで普通に見かける黄色いタンポポは、ほとんどの場合、ヨーロッパに自生し世界各地に分布を広げたセイヨウタンポポ(図1)です。セイヨウタンポポは、1904年(明治37年)には札幌において道端に生育していたという牧野富太郎博士の記録があります。このタンポポは札幌農学校(現在の北海道大学)のクラーク博士(William Smith Clark)が帰国した後、その仕事を引き継いだ農学教師ブルックス氏(William Penn Brooks)が野菜として利用するためにアメリカから導入したものが広がったと伝えられております。なお、バンクーバーオリンピックのスケート競技では、日本人選手が男女ともにメダルを獲得しましたが、ブルックス氏が札幌農学校に赴任の際にアメリカから持参したものが日本最初のスケートであるとされております。

ところで牧野博士は植物学雑誌18巻(明治37年発行)において、このタンポポは南進してやがて日本全体に広がるであろうと述べております。博士の予想どおり、今日では北海道から沖縄まで、平地だけではなく立山・室堂平や乗鞍岳・畳平などの高山帯でもセイヨウタンポを見かけるようになりました。外来タンポポには、セイヨウタンポポとともにアカミタンポポがあります。アカミタンポポはセイヨウタンポポに比べると少なく、おもに都市部やその近隣地域でセイヨウタンポポとともに生育していますが、近年、このタンポポが白山の室堂でも繁殖していることが判りました (野上2008)。白山以外の高山帯においてセイヨウタンポとともにアカミタンポポが見つかるようになるのは、それほど先のことではないと想われます。もとから生育している在来のタンポポには、研究者により認識が異なりますが15種3亜種2変種(Morita 1995)または22種4変種8品種(Kitamura 1957)が知られています。それらのなかでニホンタンポポ(カンサイタンポポ、トウカイタンポポ、カントウタンポポなどの総称)とオキタンポポは二倍体植物で、その他のタンポポは二種の外来種を含め、すべて三倍体以上の倍数体です。二倍体の個体は、私たちヒトと同様、1個の細胞内では染色体は2本ずつの相同染色体組から構成されております。二倍体個体では、減数分裂において2本の相同染色体は対合して二価染色体を形成したあとで分離しますので、正常な減数分裂により生じた花粉と卵細胞は、親個体の半数の染色体しかありません。それらの受精により生じた個体は、親個体と同数の染色体をもつことになります。しかし三倍体以上の個体では正常な減数分裂は行われません。三倍体以上の奇数倍数体では、正常な花粉や卵細胞が形成されないため、正常な種子は稔らないのが一般的です。しかし、すべてのタンポポには果実が稔ります。三倍体以上の奇数倍数体個体では正常な花粉や卵細胞が形成されないのに果実が稔るのはどうしてでしょうか。その理由は、わが国の三倍体以上の倍数体タンポポでの種子は母親の体細胞から形成されるからです。母親の体細胞から種子ができますので、発芽して成長したタンポポは、母親と全く同一の染色体・遺伝子をもっています。種子で殖えているのにもかかわらず、栄養繁殖で殖えた個体と同様、同一の遺伝子をもつクローンです。ところでタンポポの花は、一本の花茎に一個の花が咲いているように見えますが,ヒマワリ、コスモスなど他のキク科植物の花と同様に、実は小さな花が多数集まった頭状花とよばれる花が咲きます。一本の花茎のタンポポの果実は、一粒一粒がそれぞれが一個の花から稔ったものなのです。一本の花茎に母親と同一の遺伝子を持つ多数の果実ができ、やがて白い冠毛で風を受けて飛散することになります。
富山大学五福キャンパス内には、春になるとグラウンドを中心にセイヨウタンポポの花がたくさん咲きます。タンポポは一本の花茎に多数の果実をつけますので、富山大学五福キャンパス内に生えているセイヨウタンポポは、もともとは少数の個体から殖えていった可能性があります。そこで大学内のセイヨウタンポポの染色体を調べてみました。その結果、三倍体(2n=24)と四倍体(2n=32)が混ざって生えていることが判りました(図2)。
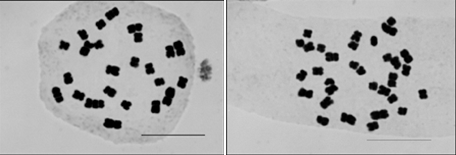
および四倍体セイヨウタンポポ(2n=32)の染色体 (右)
また富山県内の10カ所からも合わせて386個体のセイヨウタンポポを集めて調べたところ、三倍体は231個体、四倍体が155個体でした。10カ所すべてで三倍体と四倍体が見つかりましたので、三倍体と四倍体が混ざって生えるのは富山県内では一般的な状態であることが判りました。富山大学五福キャンパス内から採集した三倍体(2n=24)五株について核型を調べたところ、二株は同じ核型を示しましたが、他はすべて異なっておりました。つまり、たった五株の三倍体を調べただけで、核型の異なる四つの系統が見つかったのです。セイヨウタンポポは、通常は交配することなく母個体の体細胞から発達した果実によって殖えていきますので、形質が他の個体とは混ざり合うことはありません。従って形質の違いに基づき区別された系統のそれぞれは微細種と呼ばれております。その数は千程度知られておりますが、富山大学五福キャンパス内だけでも、かなりの数の核型の異なるセイヨウタンポポ系統(微細種)が生育しているものと想われます。
在来の倍数体タンポポにおいても、セイヨウタンポポと同様に倍数性や核型の異なる系統が複数存在することが判りました。九州や四国に多いシロバナタンポポ(図4)は五倍体であり、1クローンであると見なされておりました。しかし、染色体を調べた結果、九州には四倍体も生育すること、五倍体には核型で区別される2つのタイプが存在することが判りました。エゾタンポポは本州の中部以北および北海道に分布するタンポポです。富山県内には三倍体だけが自生しますが、日本全体には三倍体から十一倍体までの倍数体が存在することが判りました(種内倍数性)。キビシロタンポポ、ヤマザトタンポポでも倍数性が存在することが判っております。
倍数体タンポポから生じる個体は母親タンポポの体細胞に起源するクローンです。エゾタンポポやシロバナタンポポをはじめ、三倍体以上の倍数体タンポポに一般的な種内倍数性は、どのようにして形成されたのでしょうか。

倍数体タンポポと二倍体タンポポとの交配によって、新たな倍数体が形成されて倍数性が生じたとする考えがあります。この場合、ニホンタンポポやオキタンポポの染色体を倍数体タンポポは持っているはずです。しかし核型を観察する限り、持っているようには見えません。体細胞での倍数化によって染色体数が変化(突然変異)することにより倍数性が生じたことも考えられます。その場合には、二倍体が倍数化し続けると二倍体→四倍体→八倍体と変化しますし、三倍体が元になったとしたら三倍体→六倍体→十二倍体と変化するはずです。シロバナタンポポでは四、五倍体、エゾタンポポには三倍体、四倍体、五倍体、六倍体、七倍体、八倍体、九倍体、十倍体、十一倍体が存在します。それらの変化は体細胞の倍数化では説明できません。どのようなしくみでタンポポ属の倍数進化が起こったのか、解明の手がかりさえ未だ得られておりません。タンポポ属の倍数化の解明は、未だ知られていない染色体進化の新たなしくみの理解に寄与するはずです。